




摘要:【目的】研究高氧氣調(diào)包裝(HiOx,80% O2/20% CO2)對(duì)宰后豬肉蛋白質(zhì)氧化、鈣蛋白酶活性及蛋白質(zhì)降解的影響,探討高氧氣調(diào)包裝影響豬肉品質(zhì)的內(nèi)在機(jī)制。【方法】選取12條冷卻(4℃)24h后的杜洛克×長(zhǎng)白×約克夏三元雜交豬背最長(zhǎng)肌,分別進(jìn)行高氧氣調(diào)包裝和真空包裝(VP),4℃冷庫貯藏,分別在1、4、6d 測(cè)定羰基含量及分布、巰基含量、肌節(jié)變化、鈣蛋白酶活性、肌聯(lián)蛋白及肌鈣蛋白-T 降解變化。【結(jié)果】高氧氣調(diào)包裝組羰基含量高于真空包裝組且貯藏第4和6天差異顯著(P<0.05)。貯藏第1和4天,高氧氣調(diào)包裝組肌細(xì)胞外圍出現(xiàn)羰基氧化熒光信號(hào),熒光以靠近細(xì)胞膜處密度更高,并且逐漸向細(xì)胞內(nèi)部擴(kuò)散;貯藏第6天,高氧氣調(diào)包裝組細(xì)胞膜呈高亮熒光圈,胞內(nèi)熒光增強(qiáng),而真空包裝組熒光信號(hào)較弱。貯藏第6天,高氧氣調(diào)包裝組巰基含量顯著低于真空包裝組(P<0.05)。真空包裝組宰后肌節(jié)M線弱化、A帶模糊、肌原纖維Z線斷裂;高氧氣調(diào)包裝組肌節(jié)結(jié)構(gòu)相對(duì)完整。高氧氣調(diào)包裝組在貯藏第1 天鈣蛋白酶活性顯著低于真空包裝組(P<0.05);高氧氣調(diào)包裝抑制了肌聯(lián)蛋白和肌鈣蛋白-T 的降解,且在貯藏第4和6天差異顯著(P<0.05)。【結(jié)論】高氧氣調(diào)包裝能夠顯著提高宰后豬肉蛋白質(zhì)氧化程度,抑制鈣蛋白酶活性發(fā)揮及其底物蛋白質(zhì)的降解。
關(guān)鍵詞:豬肉;高氧氣調(diào)包裝;真空包裝;蛋白質(zhì)氧化;鈣蛋白酶活性;蛋白質(zhì)降解
引言
【研究意義】隨著人們生活節(jié)奏的加快,包裝銷售因其高度便捷性受到越來越多消費(fèi)者的歡迎。高氧氣調(diào)包裝、真空包裝以及透氧包裝是目前肉制品行業(yè)3大主要包裝方式。高氧氣調(diào)包裝常以80%O2和20%CO2相混合的方式對(duì)肉品進(jìn)行包裝,一方面高濃度氧氣的存在可以使冷卻肉呈現(xiàn)亮紅色,更能刺激消費(fèi)者的購(gòu)買欲;另一方面,二氧化碳能夠抑制微生物生長(zhǎng)而延長(zhǎng)商品貨架期。然而高氧氣調(diào)包裝的富氧環(huán)境不可避免會(huì)帶來蛋白質(zhì)氧化的問題,加強(qiáng)此方面的研究,對(duì)探討高氧氣調(diào)包裝影響豬肉品質(zhì)的內(nèi)在機(jī)制具有重要意義?!厩叭搜芯窟M(jìn)展】前期研究表明氧化會(huì)導(dǎo)致蛋白質(zhì)羰基化,一些必需氨基酸如精氨酸、蘇氨酸、賴氨酸結(jié)構(gòu)上會(huì)發(fā)生不可逆轉(zhuǎn)的氧化修飾,使得蛋白質(zhì)功能喪失?,F(xiàn)已證實(shí),蛋白質(zhì)氧化能夠?qū)е码逆溂肮δ芑鶊F(tuán)結(jié)構(gòu)改變、蛋白酶活性喪失、影響蛋白水解敏感性,以及促進(jìn)分子間交聯(lián)、聚集產(chǎn)生。動(dòng)物宰后肌肉成熟過程會(huì)發(fā)生一系列復(fù)雜的生理生化變化。其中,肌原纖維蛋白以及一些肌肉骨架蛋白的降解,對(duì)維持肌原纖維結(jié)構(gòu)完整性十分重要。蛋白水解會(huì)破壞肌原纖維結(jié)構(gòu)的完整性,改變肌細(xì)胞的有序結(jié)構(gòu),影響肌細(xì)胞間收縮與傳導(dǎo)過程,從而影響肌細(xì)胞內(nèi)水分含量、分布以及肉質(zhì)嫩化。而這些蛋白被認(rèn)為是鈣蛋白酶所降解的底物蛋白。宰后早期鈣蛋白酶活性的變化被認(rèn)為是調(diào)控肉品質(zhì)改善的關(guān)鍵原因之一。然而,鈣蛋白酶的活性位點(diǎn)上存在組氨酸以及含有巰基的半胱氨酸殘基,這些是極易被氧化的基團(tuán),從而使鈣蛋白酶因氧化而喪失活性。ROWE等對(duì)牛肉進(jìn)行輻照氧化處理,證實(shí)了蛋白質(zhì)氧化可通過抑制鈣蛋白酶活性來降低宰后成熟過程中蛋白質(zhì)降解程度,輻照氧化組嫩度顯著低于未輻照組,且肌間線蛋白和肌鈣蛋白-T 降解產(chǎn)物更少,鈣激活酶的自溶程度較低。然而,蛋白質(zhì)氧化調(diào)控宰后肉品品質(zhì)的具體機(jī)制尚不清楚。【本研究切入點(diǎn)】目前,在肉的成熟過程中,肌原纖維蛋白和與其降解相關(guān)的關(guān)鍵酶被氧化后,底物蛋白的結(jié)構(gòu)、降解情況變化以及關(guān)鍵酶的活性變化尚未被闡明,有關(guān)高氧氣調(diào)包裝下蛋白質(zhì)氧化影響豬肉鈣蛋白酶活性、底物蛋白質(zhì)降解等尚無報(bào)道?!緮M解決的關(guān)鍵問題】本試驗(yàn)以高氧氣調(diào)包裝和真空包裝的杜洛克×長(zhǎng)白×約克夏三元雜交豬背最長(zhǎng)肌為研究對(duì)象,從蛋白質(zhì)氧化、鈣蛋白酶活性及蛋白質(zhì)降解3方面揭示高氧氣調(diào)包裝影響宰后豬肉成熟品質(zhì)的內(nèi)在機(jī)制,為指導(dǎo)高氧氣調(diào)包裝方式的生產(chǎn)應(yīng)用提供理論依據(jù)。
1 材料與方法
試驗(yàn)于2015年在南京農(nóng)業(yè)大學(xué)食品科技學(xué)院進(jìn)行。
1.1 材料與試劑
試驗(yàn)以杜洛克×長(zhǎng)白×約克夏三元雜交商品豬背最長(zhǎng)肌為研究對(duì)象。采樣6頭6月齡、活體重為(100±5)kg健康商品豬,于江蘇淮安蘇食屠宰場(chǎng)宰殺。在4℃冷庫排酸24h,取左右背最長(zhǎng)肌共12條。
聚丙烯熱成型托盤、封口膜和真空包裝袋;DNPH(2,4-二硝基苯肼,2,4-dinitrophenylhydrazine)和DTNB(5,5’二硫代-2-硝基苯甲酸,5,5Dithiobis-2-nitrobenzoic acid);鹽酸胍;TCA(三氯乙酸,trichloroacetic acid);CY3-羊抗兔IgG(BA1032)、山羊及兔血清;DNPH 抗體(D9556)和肌鈣蛋白-T 抗體(T6277);HRP 標(biāo)記羊抗鼠IgG(ab6789);ECL化學(xué)發(fā)光試劑盒(NCI4106)和BCA 試劑盒(23250);聚丙烯酰胺凝膠電泳緩沖液(161-0732)和聚丙烯酰胺凝膠電泳轉(zhuǎn)移液(161-0734);蛋白預(yù)制凝膠
?。╓002-6);其他試劑均為市售分析純。
1.2 儀器與設(shè)備
Smart 500氣調(diào)包裝機(jī);真空包裝機(jī);H-7650透射電子顯微鏡;LSM700META 激光共聚焦顯微鏡;CM1950超薄冷凍切片機(jī);PowerPac1000垂直電泳儀和Mini-protein Tetra Cell 電泳設(shè)備;ImageQuant LAS 4000分子成像系統(tǒng)和Imagescanner凝膠成像系統(tǒng);UV-2450紫外分光光度計(jì);Spectral Max M2e酶標(biāo)儀;Ultra Turrax T25 BASIS高速勻漿機(jī);Avanti J-E 落地式高速冷凍離心機(jī);TW20水浴鍋;低溫冰柜。
1.3 方法
1.3.1 樣品包裝 剔除背最長(zhǎng)肌表面筋膜、脂肪及結(jié)締組織,均分為3塊,修剪成厚約2.5cm、重400g左右的肉塊,肉樣隨機(jī)分為兩組分別進(jìn)行高氧氣調(diào)包裝和真空包裝。高氧氣調(diào)包裝:首先將樣品放入聚丙烯熱成型托盤(6cm×23cm×13cm)中,采用填充比例為80% O2/20% CO2、充氣壓力為105kPa 的混合氣體,于140℃封口(標(biāo)準(zhǔn)大氣壓下封口膜透氧率<1 cm3·m-2·d-1)完成高氧氣調(diào)包裝。真空包裝:采用聚丙烯材質(zhì)的真空包裝袋,標(biāo)準(zhǔn)大氣壓下透氧率<30 cm3·m-2·d-1,真空包裝機(jī)抽真空至氣壓<1 kPa,抽空30 s,100℃熱封5s。兩種包裝的肉樣在4℃冷庫中貯藏,在第1、4、6天取樣進(jìn)行指標(biāo)測(cè)定。
1.3.2 肌原纖維蛋白和肌漿蛋白的制備 肌原纖維蛋白提取參考XIONG并稍作修改。將包裝成熟1、4、6 天肉樣取出,稱取1 g 剔除脂肪及筋膜的樣品,加入5倍體積的肌原纖維蛋白提取液(10 mmol·L-1 磷酸鹽緩沖液、100 mmol·L-1 氯化鈉、2 mmol·L-1 氯化鎂、1 mmol·L-1 EGTA,pH 7.0)。4℃勻漿30s,10s間隔一次,轉(zhuǎn)速為15 000 r/min。勻漿液以1 000×g轉(zhuǎn)速離心10min,取沉淀。以上步驟重復(fù)3 次。離心后沉淀中加入5倍體積的100 mmol·L-1氯化鈉洗脫液。15000 r/min 轉(zhuǎn)速勻漿30s,10s 間隔一次。勻漿液以1000×g 轉(zhuǎn)速離心10 min,取沉淀,以上步驟重復(fù)3次。離心后所得沉淀即為肌原纖維蛋白。肌原纖維蛋白與電泳上樣緩沖液(125 mmol·L-1 Tris-base、4%SDS、20%甘油、0.5%β-巰基乙醇、0.1%溴酚藍(lán),pH6.8)充分混合,BCA試劑盒法測(cè)定肌原纖維蛋白濃度后統(tǒng)一蛋白質(zhì)濃度為7 mg·mL-1。蛋白質(zhì)溶液按體積比1:1與電泳上樣緩沖液混合,50℃水浴20 min。分裝樣品,-80℃存放用于蛋白電泳檢測(cè)。
肌漿蛋白的提取參考VEISETH 等的方法并稍作修改。將包裝后成熟1d的肉樣取出,稱取1g剔除筋膜和脂肪的肉樣,加入3倍體積4℃預(yù)冷的活性蛋白提取液(100 mmol·L-1 Tris-base、0.1%β-巰基乙醇、10 mmol·L-1 EDTA,pH 8.3),15 000 r/min 勻漿2次,每次15s,5s 間隔一次。勻漿液以轉(zhuǎn)速15000×g,4℃離心30min,所得上清通過濾紙過濾后,BCA試劑盒法測(cè)定蛋白質(zhì)濃度。去離子水調(diào)節(jié)統(tǒng)一蛋白質(zhì)濃度為8mg·mL-1,按蛋白體積﹕活性電泳上樣緩沖液(150 mmol·L-1 Tris-base、20%甘油、0.75%β-巰基乙醇、0.02%溴酚藍(lán),pH 6.8)=1﹕1 進(jìn)行稀釋。混合后樣品進(jìn)行分裝,-80℃冷凍用于鈣蛋白酶活性檢測(cè)。
1.3.3 羰基含量測(cè)定 參考ZHANG等的方法并稍作修改。稱取0.5g剔除脂肪及筋膜的肉樣,加入5mL焦磷酸鹽緩沖液勻漿3次,每次10s,轉(zhuǎn)速最高為15000 r/min。取0.5 mL勻漿液置于2個(gè)2mL離心管中,加入0.5mL 20%三氯乙酸(TCA),振蕩混勻后以12000×g離心5min。除去上清液,沉淀加入0.5mL10% TCA 振蕩混勻后再次離心,離心條件同上。對(duì)照組將離心后沉淀加0.5mL2mol·L-1HCl,試驗(yàn)組加0.5mL 10mmol·L-1 2,4-二硝基苯肼(DNPH)。暗處理30min,每隔10min迅速取出混勻10 s。待DNPH 與羰基充分反應(yīng),對(duì)照組與試驗(yàn)組加入0.5 mL 20% TCA,振蕩混勻后相同條件離心。沉淀用10mmol·L-1 HCl(溶解于1﹕1(vol/vol)乙醇/乙酸乙酯)重復(fù)洗脫3次。所得沉淀加入1mL 6mol·L-1鹽酸胍(溶解在20 mmol·L-1磷酸二氫鉀溶液,pH 2.3),4℃震蕩過夜。未溶解物12 000×g 離心5min,上清液在紫外370nm 處測(cè)定吸光度。消光系數(shù)為22000L·mol-1·cm-1,羰基含量用每毫克蛋白里含有羰基的納摩爾數(shù)表示。
1.3.4 羰基肌細(xì)胞內(nèi)分布測(cè)定 采用免疫熒光法,參考ASTRUC 等的方法并做一定修改。將包裝后成熟1、4、6d 的肉樣取出,隨機(jī)選取肉樣區(qū)域沿著平行于肌原纖維方向?qū)⒓∪馇谐扇鈼l并立即浸泡于液氮中30s,每個(gè)肉樣取6個(gè)平行。冰凍超薄切片機(jī)切取10μm 厚切片,37℃烘干2h 后與0.04%DNPH(溶解在20 mmol·L-1 磷酸鹽和0.1 mol·L-1 氯化鈉的混合液中,pH 6.0)暗處反應(yīng)16h。反應(yīng)結(jié)束后樣品用含0.1%吐溫的PBS(pH 6.75)重復(fù)沖洗(5 min,6次)。37℃兔血清一次封閉切片1h。去除封閉液,滴加1﹕100稀釋DNPH 多克隆抗體孵育,4℃反應(yīng)過夜。一抗孵育結(jié)束后,切片樣品用含0.1%吐溫的PBS(pH 6.75)重復(fù)沖洗(5min,6次),37℃山羊血清二次封閉1h。去除封閉液,滴加CY3標(biāo)記的羊抗兔免疫球蛋白溶液(1﹕50)避光孵育1h,PBS漂洗(5min,6次)。采用激光共聚焦顯微鏡在激發(fā)波長(zhǎng)555 nm、發(fā)射波長(zhǎng)570 nm 下,將樣品放大200 倍,統(tǒng)一曝光時(shí)間下觀察羰基在肌細(xì)胞內(nèi)分布。
1.3.5 巰基含量測(cè)定 巰基測(cè)定參考ELLMAN的方法,并稍作修改。0.5mL 肌原纖維蛋白溶液(4mg·mL-1)加入4.5mL 緩沖液(50 mmol·L-1磷酸鹽緩沖液、10 mmol·L-1 EDTA、0.6 mol·L-1氯化鉀,pH7.0),與0.5 mL 10 mmol·L-1 5、5-二硫代-2-硝基苯甲酸(0.2 mol·L-1磷酸鹽緩沖液配置,pH 8.0)混合均勻。混合液在40℃水浴中反應(yīng)25 min,412nm 處測(cè)定吸光度值。摩爾吸光系數(shù)為136 000 L·mol-1·cm-1,活性巰基含量用每毫克蛋白里含有巰基的納摩爾數(shù)表示。
1.3.6 電鏡觀察肌節(jié)變化 參考徐舶等方法,將成熟1、4、6d的樣品沿著肌原纖維方向切成約0.5cm×0.5 cm×0.5 cm的肉條,肉條浸泡在2.5%戊二醛中(20 mmol·L-1磷酸緩沖液溶解,pH 7.2)初步固定7d,每隔一天更新戊二醛固定液。樣品用2%餓酸二次固定,固定完成后經(jīng)磷酸緩沖液反復(fù)沖洗,梯度乙醇脫水,100%丙酮置換,最后采用Spurr 樹脂浸透包埋。超薄切片機(jī)進(jìn)行切片,日立H-7650 透射顯微鏡將樣本放大3000倍觀察肌節(jié)變化。
1.3.7 鈣蛋白酶活性測(cè)定 參考酪蛋白酶原法,采用4%的濃縮膠(丙烯酰胺﹕甲叉丙烯酰胺=75﹕1,125 mmol·L-1 Tris-base(pH 6.8),0.05%過硫酸銨,0.05%四甲基乙二胺)和12.5%的分離膠(丙烯酰胺﹕甲叉丙烯酰胺=75﹕1,375 mmol·L-1 Tris-base(pH8.8),2.1 mmol·L-1 酪蛋白,0.05%過硫酸銨,0.05% 四甲基乙二胺)。緩慢加入4℃預(yù)冷的電泳緩沖液(25mmol·L-1 Tris-base,192 mmol·L-1 甘氨酸,1mmol·L-1EDTA,0.05%β-巰基乙醇),接通電源100V,預(yù)跑15min 后上樣。蛋白樣品在100V、4℃環(huán)境電泳6h至電泳結(jié)束?;钚噪娪痉磻?yīng)液(50 mmol·L-1 Tris-base、5 mmol·L-1 氯化鈣、0.1%β-巰基乙醇,pH 7.8)室溫孵育膠體前1 h,每20 min 更換活性電泳反應(yīng)液。繼續(xù)孵育16 h 后膠體進(jìn)行染色和脫色,凝膠成像儀拍照。鈣蛋白酶活性條帶采用Quanity one 分析軟件進(jìn)行半定量分析。
1.3.8 肌聯(lián)蛋白和肌鈣蛋白-T 降解測(cè)定 肌聯(lián)蛋白采用5%的連續(xù)聚丙烯酰胺連續(xù)電泳(丙烯酰胺﹕甲叉丙烯酰胺=100﹕1,1.5 mol·L-1 Tris-base(pH 8.0),0.1% SDS,0.05%過硫酸銨,0.05%四甲基乙二胺)。每塊膠體恒流4.5mA電泳20h至電泳結(jié)束。肌聯(lián)蛋白膠體采用改進(jìn)型考馬斯亮藍(lán)染色和脫色。肌鈣蛋白-T 的聚丙烯酰胺變性凝膠電泳采用濃縮膠濃度4%、分離膠濃度10%的蛋白預(yù)制膠。恒壓80V電泳30min后,調(diào)整電壓為120V至溴酚藍(lán)標(biāo)記跑至膠體底部,電泳結(jié)束。取出肌鈣蛋白-T 的膠體通過濕法轉(zhuǎn)印技術(shù)將蛋白轉(zhuǎn)移到聚偏氟乙烯膜上。轉(zhuǎn)印條件為恒壓70V,轉(zhuǎn)印70min。轉(zhuǎn)印完成后將聚偏氟乙烯膜正面放入含有5%牛血清白蛋白的TBST(20mmol·L-1 Tris-base,137 mmol·L-1 氯化鈉,5 mmol·L-1氯化鉀,0.05%吐溫20)溶解液中,室溫封閉2 h。肌鈣蛋白-T 抗體(1﹕500)4℃繼續(xù)孵育16 h。TBST 反復(fù)漂洗(5min,6次)后將聚偏氟乙烯膜置于HRP標(biāo)記二抗(1﹕500)中,室溫反應(yīng)2h。TBST 反復(fù)漂洗(5min,6次),膜表面滴加化學(xué)顯色劑ECL,暗箱反應(yīng)6min后終止反應(yīng),凝膠成像儀掃描并拍照。蛋白免疫印記條帶采用Quanity one 分析軟件進(jìn)行半定量分析。
1.4 數(shù)據(jù)分析
數(shù)據(jù)采用SAS 8.1 統(tǒng)計(jì)軟件進(jìn)行方差分析,并通過Duncan’ 法比較處理組間差異,差異顯著性水平為P<0.05,數(shù)據(jù)以均值±標(biāo)準(zhǔn)差(means±SD,n=6)表示。
2 結(jié)果
2.1 高氧氣調(diào)包裝對(duì)豬肉羰基含量及肌細(xì)胞內(nèi)分布的影響
圖1 結(jié)果顯示,貯藏第1 天處理組間羰基含量無顯著差異(P>0.05),貯藏第4 天和第6 天,高氧氣調(diào)包裝組羰基含量顯著高于真空包裝組(P<0.05)。
激光共聚焦顯微鏡觀察結(jié)果見圖2,其中陰性對(duì)照組肌細(xì)胞形態(tài)比較規(guī)則,胞膜完整,細(xì)胞飽滿,細(xì)胞間隙致密,無熒光著色。圖示紅色熒光信號(hào)部分即特異性結(jié)合的氧化羰基。高氧氣調(diào)包裝組和真空包裝組熒光信號(hào)在貯藏期間均呈現(xiàn)動(dòng)態(tài)變化。宰后貯藏第1天,高氧氣調(diào)包裝組視野下部分肌細(xì)胞外周近胞膜處出現(xiàn)氧化熒光信號(hào),熒光分布不均勻。宰后貯藏第4天,高氧氣調(diào)包裝組熒光多分布于細(xì)胞膜邊緣,以細(xì)胞靠膜處密度更高,逐漸向胞內(nèi)擴(kuò)散呈暗紅色。真空包裝組熒光分布于細(xì)胞膜包圍的很小區(qū)域,熒光信號(hào)較弱。宰后貯藏第6天,高氧氣調(diào)包裝組肌細(xì)胞外周呈高亮熒光圈,細(xì)胞內(nèi)部熒光信號(hào)增強(qiáng)。肌細(xì)胞間熒光信號(hào)分布基本均勻。此外,相對(duì)于高氧氣調(diào)包裝組,真空包裝組熒光信號(hào)較弱,分布多集中在細(xì)胞膜周圍,且肌細(xì)胞變形嚴(yán)重,肌細(xì)胞形態(tài)出現(xiàn)脫水、皺縮,骨架結(jié)構(gòu)坍塌,細(xì)胞間裂隙變大。

同天平均值數(shù)據(jù)上標(biāo)有不同小寫字母者表示差異顯著(P<0.05)。下同
圖1 高氧氣調(diào)包裝對(duì)宰后貯藏期間豬肉羰基含量的影響
A:陰性對(duì)照;B、C、D 分別對(duì)應(yīng)宰后貯藏1、4、6 d 時(shí)高氧氣調(diào)包裝豬肉肌細(xì)胞內(nèi)羰基分布;E:陰性對(duì)照(明場(chǎng));F、G、H 分別對(duì)應(yīng)宰后貯藏1、4、6 d 時(shí)真空包裝豬肉肌細(xì)胞內(nèi)羰基分布;視野放大倍數(shù)均為200 倍;標(biāo)尺表示100 μm

圖2 高氧氣調(diào)包裝和真空包裝豬肉肌細(xì)胞內(nèi)羰基分布的免疫熒光顯微鏡
2.2 高氧氣調(diào)包裝對(duì)豬肉活性巰基含量的影響
由圖3可知,宰后貯藏期間高氧氣調(diào)包裝組的活
性巰基含量均低于真空包裝組。在貯藏第6 天,高氧氣調(diào)包裝組活性巰基含量與真空包裝組差異顯著(P<0.05)。

圖3 高氧氣調(diào)包裝對(duì)宰后豬肉活性巰基含量的影響
2.3 高氧氣調(diào)包裝對(duì)豬肉肌節(jié)的影響
由圖4可知,高氧氣調(diào)包裝和真空包裝組肌原纖維超微結(jié)構(gòu)在宰后貯藏期間均發(fā)生顯著變化。宰后貯藏第1天,高氧氣調(diào)包裝組和真空包裝組A帶和I帶清晰可辨,Z線和M 線均保持完整,肌原纖維之間連接緊密,處理組間無明顯差異。宰后貯藏第4天,相對(duì)于高氧氣調(diào)包裝組,真空包裝組Z線附近發(fā)生較為明顯斷裂,Z線周圍的細(xì)絲與Z線的連接開始弱化,M線也出現(xiàn)弱化。宰后貯藏第6天,真空包裝相對(duì)于高氧氣調(diào)包裝,大量肌節(jié)出現(xiàn)Z線的裂解,肌節(jié)分裂成細(xì)絲狀,A帶模糊,M線基本消失,肌節(jié)結(jié)構(gòu)遭到破壞。肌節(jié)形態(tài)學(xué)結(jié)果表明,高氧氣調(diào)包裝抑制了宰后豬肉成熟過程。

A、B、C 分別對(duì)應(yīng)宰后貯藏1、4、6 d 時(shí)高氧氣調(diào)包裝豬肉肌節(jié)變化;D、E、F 分別對(duì)應(yīng)宰后貯藏1、4、6 d 時(shí)真空包裝豬肉肌節(jié)變化;視野放大倍數(shù)×3000 倍;標(biāo)尺表示1.0
圖4 高氧氣調(diào)包裝和真空包裝宰后豬肉肌節(jié)變化的透射電鏡圖
2.4 高氧氣調(diào)包裝對(duì)豬肉鈣蛋白酶活性的影響
從圖5和表1可知,高氧氣調(diào)包裝組在宰后貯藏第1天μ-鈣蛋白酶條帶相對(duì)光密度值顯著高于真空包裝組(P<0.05),即相對(duì)于真空包裝組,高氧氣調(diào)包裝組μ-鈣蛋白酶活性顯著低于真空包裝組(P<0.05),高氧氣調(diào)包裝組豬肉在宰后早期μ-鈣蛋白酶活力的發(fā)揮受到抑制。
2.5 高氧氣調(diào)包裝對(duì)豬肉蛋白質(zhì)降解的影響
2.5.1 高氧氣調(diào)包裝對(duì)豬肉肌聯(lián)蛋白降解的影響 由圖6和表2可知,在宰后貯藏1d,高氧氣調(diào)包裝組和真空包裝組降解條帶含量沒有明顯差異(P>0.05)。在宰后貯藏第4 和6 天,真空包裝組肌聯(lián)蛋白降解條帶含量顯著低于高氧氣調(diào)包裝組(P<0.05),即高氧氣調(diào)包裝組抑制了宰后豬肉肌聯(lián)蛋白的降解。

圖5 高氧氣調(diào)包裝對(duì)貯藏1 天豬肉鈣蛋白酶活性的影響
表1 高氧氣調(diào)包裝和真空包裝豬肉宰后貯藏第1 天鈣蛋白酶活性
| 指標(biāo) | 貯藏時(shí)間 | 高氧氣調(diào)包裝機(jī) | 真空包裝 |
| μ-鈣蛋白酶 | 1 | 3.22±0.15a | 0.98±0.03b |
鈣蛋白酶相對(duì)于標(biāo)準(zhǔn)樣品的光密度值;不同小寫字母者表示差異顯著(P<0.05)
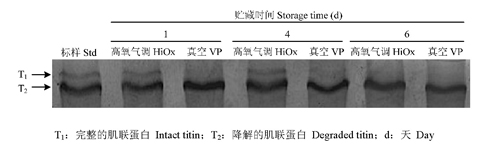
T1:完整的肌聯(lián)蛋白 ;T2:降解的肌聯(lián)蛋白;d:天
圖6 高氧氣調(diào)包裝對(duì)豬肉肌聯(lián)蛋白降解的影響
表2 高氧氣調(diào)包裝和真空包裝豬肉肌聯(lián)蛋白蛋白質(zhì)降解的變化
| 指標(biāo) | 貯藏時(shí)間 | 高氧氣調(diào)包裝 | 真空包裝 |
| 降解的肌聯(lián)蛋白(T2) | 1 | 1.04±0.08a | 1.08±0.08a |
| Degraded(T2) | 4 | 1.02±0.02a | 0.90±0.01b |
| 6 | 0.57±0.01a | 0.48±0.02b |
降解的肌聯(lián)蛋白相對(duì)于標(biāo)準(zhǔn)樣品的光密度值;同天不同小寫字母表示差異顯著(P<0.05)
2.5.2 高氧氣調(diào)包裝對(duì)豬肉肌鈣蛋白-T 降解的影響 由圖7和表3可知,在貯藏第4天和第6 天,高氧氣調(diào)包裝組中肌鈣蛋白-T 30 kDa 降解產(chǎn)物含量顯著低于真空包裝組(P<0.05),而貯藏第1天,高氧氣調(diào)包裝對(duì)肌鈣蛋白-T 30kDa 的降解產(chǎn)物含量并無顯著影響(P>0.05)。

圖7 高氧氣調(diào)包裝對(duì)宰后豬肉貯藏期間肌鈣蛋白-T 降解的影響
表3 高氧氣調(diào)包裝和真空包裝豬肉肌鈣蛋白-T 的降解變化
| 指標(biāo) | 貯藏時(shí)間 | 高氧氣調(diào)包裝 | 真空包裝 |
| 降解的肌鈣蛋白-T | 1 | 0.99±0.04a | 0.95±0.06a |
| Degraded troponin-T(30KDa) | 4 | 1.53±0.05a | 2.44±0.03b |
| 6 | 2.05±0.05a | 2.96±0.03b |
降解的肌鈣蛋白-T 相對(duì)于標(biāo)準(zhǔn)樣品的光密度值;同天平均值數(shù)據(jù)標(biāo)有不同小寫字母表示差異顯著(P<0.05)
3 討論
蛋白質(zhì)氧化指由氧分壓產(chǎn)生的活性氧(ROS)與蛋白質(zhì)反應(yīng)導(dǎo)致蛋白共價(jià)修飾。肌肉蛋白質(zhì)在ROS作用下,某些特定氧基酸殘基發(fā)生反應(yīng),導(dǎo)致蛋白質(zhì)結(jié)構(gòu)變化,使得蛋白質(zhì)與氧化物親和力加強(qiáng),易于產(chǎn)生水解、交聯(lián)、聚合,從而損害肌細(xì)胞功能。蛋白質(zhì)氧化損傷反應(yīng)涉及蛋白質(zhì)羰基的產(chǎn)生,羰基生成是一個(gè)復(fù)雜的過程。STADTMAN認(rèn)為羰基生成是由活性氧攻擊氨基酸自由氨基或亞氨基,經(jīng)反應(yīng)最終生成NH3 和相應(yīng)羰基衍生物。另外,ROS所致蛋白質(zhì)肽鏈斷裂,在斷裂處也可產(chǎn)生羰基。ZAKTYS-WALIWANDER 等發(fā)現(xiàn)高氧氣調(diào)包裝的牛肉在4℃貯藏8d和14d的羰基含量均顯著高于真空包裝組,同時(shí)蛋白質(zhì)氧化能夠抑制牛肉的嫩化。DELLES 等同樣發(fā)現(xiàn)高氧氣調(diào)包裝相對(duì)于真空包裝和透氧包裝,顯著增加了豬肉蛋白質(zhì)羰基含量,且證實(shí)了蛋白質(zhì)氧化是解釋宰后豬肉較差持水力的原因之一。這與本研究結(jié)果一致,由羰基含量反映的肽鏈、氨基酸結(jié)構(gòu)和功能的改變、蛋白質(zhì)氧化程度增加提示了影響宰后成熟肉品質(zhì)的可能原因。ASTRUC 等認(rèn)為,相對(duì)于通過羰基含量評(píng)價(jià)蛋白質(zhì)氧化程度,通過激光共聚焦顯微鏡觀察羰基在肌細(xì)胞內(nèi)的熒光分布,更能清楚地了解肌細(xì)胞氧化通路的動(dòng)態(tài)變化。本試驗(yàn)中,高氧氣調(diào)包裝組比真空包裝組較先出現(xiàn)蛋白質(zhì)氧化,且氧化熒光信號(hào)集中于肌細(xì)胞膜邊緣,逐漸向細(xì)胞內(nèi)部擴(kuò)散,肌細(xì)胞間熒光信號(hào)分布相對(duì)均勻。ASTRUC試驗(yàn)發(fā)現(xiàn)宰后動(dòng)物體內(nèi)蛋白質(zhì)氧化起始于細(xì)胞膜;這主要因?yàn)榧?xì)胞膜是磷脂雙分子層,其富含的多不飽和脂肪酸(可保持細(xì)胞膜的相對(duì)流動(dòng)性以保證細(xì)胞正常生理功能)極易被氧化,膜上蛋白質(zhì)又因接近磷脂雙分子層,由脂肪氧化形成的羥自由基能夠奪取蛋白質(zhì)分子的氫離子,使得細(xì)胞膜上蛋白質(zhì)開始了與脂肪氧化類似的自由基鏈?zhǔn)椒磻?yīng)。ROS從肌細(xì)胞膜向細(xì)胞內(nèi)部傳遞的過程中肌漿蛋白和肌原纖維蛋白逐漸氧化。
巰基是蛋白質(zhì)殘基中最具有反應(yīng)活性的基團(tuán),極易被氧化為二硫鍵、次磺酸、亞磺酸及磺酸,或被一氧化氮亞硝基化。LUND 等通過對(duì)肌球蛋白的重鏈進(jìn)行一維電泳時(shí)發(fā)現(xiàn),蛋白質(zhì)分子間通過二硫鍵交聯(lián)、聚集,是巰基含量降低的主要原因之一。此外,目前已鑒定了一大批易受氧化還原調(diào)控的蛋白質(zhì),其活性部位均含有巰基基團(tuán),通過對(duì)巰基的修飾可以顯著改變細(xì)胞內(nèi)的信號(hào)轉(zhuǎn)導(dǎo)。HUFF-LONERGAN 等報(bào)道,鈣蛋白酶通過降解對(duì)細(xì)胞有序性和完整性十分重要的肌原纖維蛋白和細(xì)胞骨架蛋白從而調(diào)控牛肉嫩度和豬肉的保水性。然而鈣蛋白酶活性區(qū)域的巰基極易被氧化形成二硫鍵,導(dǎo)致鈣激活酶失活,底物肌原纖維蛋白降解受到抑制,肌肉細(xì)胞的有序性和完整性遭到破壞。因此,蛋白質(zhì)巰基完全可以作為影響細(xì)胞內(nèi)氧化還原信號(hào)轉(zhuǎn)導(dǎo)過程的更直接、更相關(guān)的“探針”。本研究發(fā)現(xiàn),由活性巰基含量降低反映的高氧氣調(diào)包裝下氧化程度增加,與羰基的結(jié)果相一致。
在宰后成熟過程中,蛋白質(zhì)氧化反應(yīng)主要包括肌原纖維蛋白和相關(guān)酶系統(tǒng)氧化,這些氧化都是通過改變肌原纖維的降解從而影響肉品品質(zhì)。其中,鈣蛋白酶能夠調(diào)控肌原纖維蛋白以及一些肌肉骨架蛋白的降解,例如肌聯(lián)蛋白、伴肌動(dòng)蛋白、肌鈣蛋白-T、整聯(lián)蛋白、肌間線蛋白等。目前普遍認(rèn)為肉類在宰后成熟過程中成熟品質(zhì)的改善主要?dú)w功于這些蛋白質(zhì)的降解。有關(guān)鈣蛋白酶系統(tǒng)動(dòng)力學(xué)研究發(fā)現(xiàn),氧化能改變?cè)缀髣?dòng)物體內(nèi)的離子水平,使得鈣蛋白酶活性改變,
從而導(dǎo)致底物降解情況發(fā)生改變。此外,鈣蛋白酶活性的發(fā)揮必須保證其活性區(qū)域還原性。LAMETSCH等發(fā)現(xiàn)μ-鈣蛋白酶半胱氨酸活性區(qū)域?qū)ρ趸h(huán)境十分敏感,鈣激活酶肽段殘基92—114間存在二級(jí)結(jié)構(gòu)螺旋,其可伸展性縮短Cys 108與Cys 115空間距離,從而為活性區(qū)域Cys 108 和Cys 115 之間形成二硫鍵提供可能,導(dǎo)致鈣蛋白酶活性喪失。本研究表明,由羰基含量及分布、巰基含量、肌節(jié)變化反映的蛋白質(zhì)氧化增加,鈣蛋白酶活性區(qū)域極易受到氧化攻擊,蛋白質(zhì)結(jié)構(gòu)破壞,從而抑制了鈣蛋白酶功能活性發(fā)揮。
μ-鈣蛋白酶活力可以預(yù)測(cè)宰后成熟過程中肌原纖維蛋白以及肌肉骨架蛋白的降解。不論是μ-鈣蛋白酶被氧化還是其底物蛋白被氧化,均會(huì)對(duì)肉類宰后成熟品質(zhì)產(chǎn)生影響。肌聯(lián)蛋白(titin)是肌細(xì)胞骨架蛋白中含量最多且分子量最大的蛋白,位于M 線和Z線之間,其C末端與M 線相接,N末端是Z線的組成部分,幾乎橫跨了半個(gè)肌節(jié)。titin 將粗絲維持在肌節(jié)的中央,具有保持粗絲穩(wěn)定、調(diào)節(jié)粗絲收縮、保持肌節(jié)及肌細(xì)胞完整性等功能。肌鈣蛋白-T(troponin-T)是肌鈣蛋白和原肌球蛋白相結(jié)合亞基,對(duì)鈣離子具有較高的敏感性,它能與原肌球蛋白結(jié)合調(diào)節(jié)橫紋肌的收縮。肌肉宰后成熟過程中,troponin-T由分子量35 kDa產(chǎn)生28—30 kDa的特征降解產(chǎn)物。目前,troponin-T 的降解特征產(chǎn)物已作為預(yù)測(cè)肉品成熟品質(zhì)的重要指標(biāo)之一。FU 等報(bào)道,高氧氣調(diào)包裝的宰后成熟第4—7天,牛肉的μ-鈣蛋白酶活性顯著低于真空包裝和透氧包裝組,且troponin-T的降解受到抑制。本試驗(yàn)也證實(shí)高氧氣調(diào)包裝能夠通過抑制宰后豬肉早期μ-鈣蛋白酶活性的發(fā)揮,對(duì)troponin-T 和titin 在宰后貯藏第4天和第6 天的降解具有顯著抑制作用。肌原纖維蛋白降解弱化一定程度上也解釋了高氧氣調(diào)包裝下宰后肌節(jié)超微形態(tài)的改變。肌節(jié)是組成肌原纖維的基本單位,高氧氣調(diào)包裝能夠抑制豬肉肌原纖維蛋白及細(xì)胞骨架蛋白的降解,體現(xiàn)在連接Z線和M線的titin 降解產(chǎn)物變少,構(gòu)成骨骼肌完整細(xì)絲的troponin-T 降解弱化,進(jìn)而抑制Z線附近斷裂,維持Z 線周圍細(xì)絲與Z線間的連接,阻礙肌原纖維經(jīng)宰后成熟過程中的小片化,保持肌節(jié)結(jié)構(gòu)的相對(duì)完整性。
4 結(jié)論
高氧氣調(diào)包裝能夠增加宰后豬肉羰基含量,擴(kuò)大羰基在肌細(xì)胞內(nèi)分布,降低活性巰基含量,加劇宰后豬肉蛋白質(zhì)氧化。同時(shí)高氧氣調(diào)包裝具有抑制早期μ-鈣蛋白酶活性發(fā)揮,抑制肌聯(lián)蛋白和肌鈣蛋白-T 等關(guān)鍵肌原纖維蛋白的降解,阻礙宰后豬肉肌節(jié)成熟的作用。

用")

